La vision des abeilles

Introduction à la vision abeilles
La vision des couleurs est définie comme la capacité à distinguer les surfaces colorées en fonction de leurs différents contenus chromatiques, indépendamment des différences d’intensité.
Les humains et de nombreuses espèces animales voient le monde en couleurs, ce qui offre des avantages uniques pour détecter les sources de nourriture, les partenaires et les prédateurs, entre autres
Quelle est la perception des couleurs par les abeilles ?
Les abeilles, en tant que pollinisateurs, sont attirées par les couleurs des fleurs, mais leur perception des couleurs diffère de celle des humains.
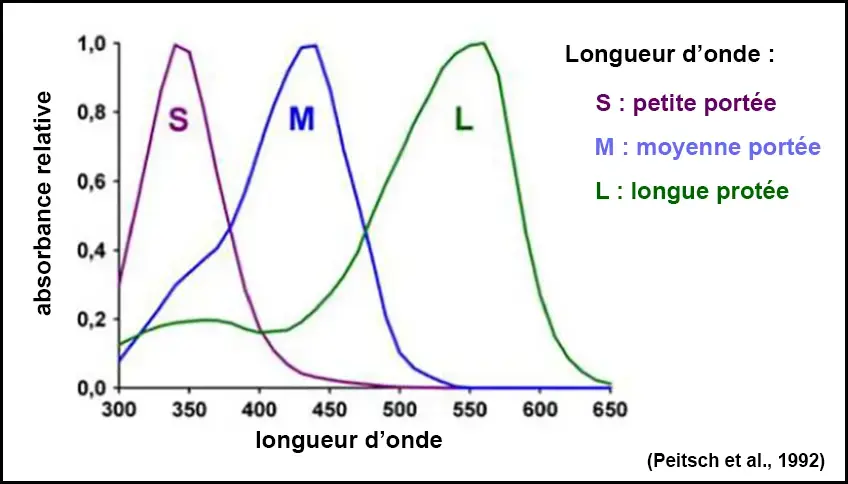
Les abeilles ont une vision trichromatique, sensible aux longueurs d’onde verte, bleue et ultraviolette, contrairement aux humains qui ont une vision trichromatique rouge, verte et bleue.
La perception des couleurs par les abeilles repose sur les trois types de photorécepteurs de leurs yeux composés.
Chaque type étant sensible à une gamme de longueurs d’onde spécifique : ultraviolet, bleu et vert, leur permettant de percevoir un spectre lumineux qui s’étend de l’ultraviolet à l’orange-rougeâtre (mais excluant le rouge).
Pour étudier comment les abeilles perçoivent les couleurs des fleurs, les chercheurs utilisent un modèle appelé « l’hexagone des abeilles » développé par Chittka (1992) qui représente l’espace chromatique des abeilles.
Ce modèle permet de classifier les couleurs des fleurs en six catégories en fonction de la façon dont les abeilles les perçoivent :
- Bleu abeille,
- Vert abeille,
- UV abeille,
- Bleu-vert abeille,
- UV-vert abeille,
- UV-bleu abeille.
L'adaptation des fleurs à la vision des abeilles
L’étude menée dans le cerrado brésilien, une savane tropicale, a révélé des informations intéressantes sur les motifs de couleurs des fleurs et leur lien avec les abeilles :
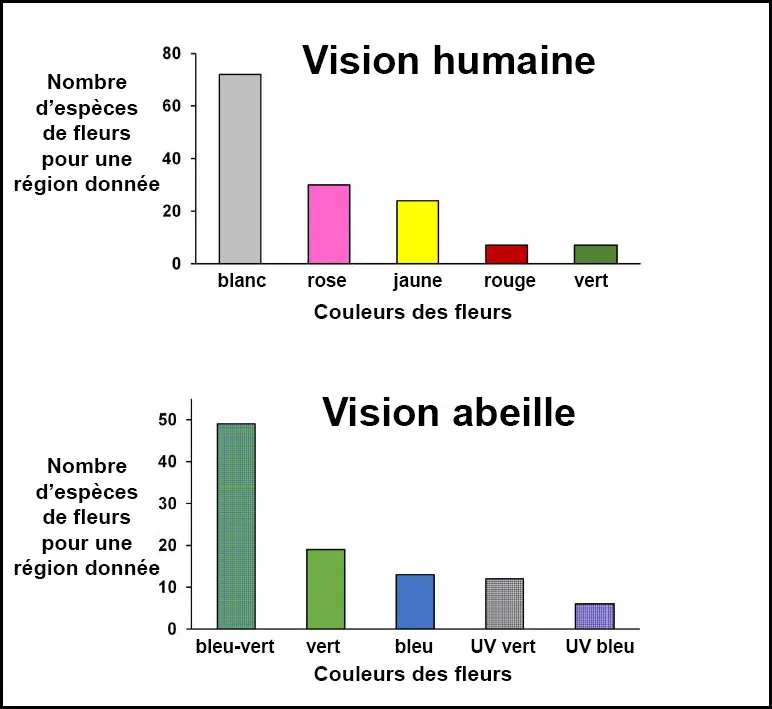
- Le bleu-vert abeille était la couleur de fleur la plus courante selon la vision des abeilles, ce qui correspond aux fleurs blanches ou blanchâtres perçues par les humains ;
- Les fleurs blanches et jaunes étaient prédominantes, suivies des fleurs roses.
Les fleurs blanches atteignaient leur pic de floraison à la fin de la saison sèche, tandis que les fleurs roses étaient plus fréquentes pendant la saison humide ; - La floraison était observée tout au long de l’année, avec une plus grande diversité de couleurs et d’espèces en fleurs à la fin de la saison sèche, période qui coïncide avec une activité accrue des abeilles ;
- Les contrastes de couleur des fleurs, importants pour leur détection par les abeilles, étaient similaires entre les saisons sèches et humides.
Cependant, le fond jaunâtre de la saison sèche, dû à la sénescence des feuilles, augmentait le contraste des couleurs des fleurs, ce qui pourrait maximiser leur détection pendant une période où l’activité florale est réduite.
En revanche, le fond plus vert de la saison humide offrait un contraste vert plus élevé, ce qui pourrait aider à la détection des fleurs parmi les feuilles vertes.
En conclusion, les motifs de couleur des fleurs dans le cerrado semblent être liés à la fois à :
- La saisonnalité,
- L’activité des abeilles,
- Leur système visuel unique.
La vision des abeilles

Les ocelles et les yeux composés chez les insectes fonctionnent comme deux systèmes visuels distincts, mais complémentaires, qui contribuent à la stabilité du vol.
Les informations des deux systèmes sont combinées dans le cerveau de l’insecte pour contrôler les réflexes de stabilisation.
Les yeux composés des abeilles fournissent des informations précises sur l’orientation et les mouvements lents, tandis que les ocelles des abeilles fournissent des informations rapides sur les changements d’orientation soudains.
Cette intégration permet à l’abeille de maintenir son équilibre et sa stabilité en vol, en ajustant rapidement sa trajectoire en réponse aux perturbations.
Vision et apprentissage visuel des abeilles
Les opsines, sont des protéines présentes dans les photorécepteurs.
Chaque type d’opsine est sensible à une gamme de longueurs d’onde spécifique, ce qui permet aux abeilles de distinguer les couleurs.
Des études se concentrant sur deux opsines sensibles au vert, Amlop1 et Amlop2, révèlent une ségrégation stricte de ces deux opsines dans le système visuel de l’abeille :
- Amlop1 est principalement présente dans les ommatidies des yeux composés et joue un rôle crucial dans la vision chromatique et l’apprentissage visuel des couleurs.
Amlop2 se trouve exclusivement dans les ocelles, un groupe de trois yeux simples situés sur le dessus de la tête de l’abeille.
Les études confirment le rôle crucial de Amlop1 dans la vision des couleurs et l’apprentissage visuel, et suggèrent que les ocelles et l’opsine Amlop2 ne sont pas impliqués dans ces processus.
Les yeux composés des abeilles
Les yeux composés sont capables de percevoir les couleurs grâce à la présence de différents types de photorécepteurs sensibles à des longueurs d’onde spécifiques.
La structure des yeux composée des abeilles, est composés d’ommatidies, les unités fonctionnelles de l’œil.
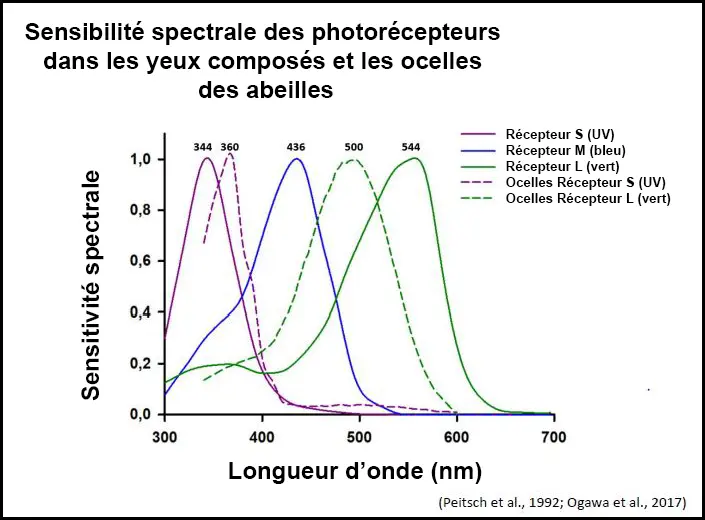
Chaque ommatidie contient neuf photorécepteurs disposés de manière concentrique.
La répartition des différents types de photorécepteurs dans les ommatidies n’est pas uniforme, il existe trois types d’ommatidies (type I, II et III) avec des proportions variables de photorécepteurs sensibles à l’ultraviolet, au bleu et au vert.
Les yeux composés des abeilles, avec leur structure complexe et leur champ visuel large, excellent dans la détection des mouvements lents et des détails de l’environnement.
Ils sont essentiels pour l’orientation spatiale générale et la perception du mouvement relatif à l’environnement.
Voici les principaux points à retenir sur les yeux composés des abeilles :
- Structure modulaire : Les yeux composés sont constitués de milliers d’unités fonctionnelles appelées ommatidies. Chaque ommatidie possède sa propre lentille et son propre groupe de photorécepteurs, fonctionnant comme un mini-œil.
- Diversité des photorécepteurs : Les abeilles possèdent trois types de photorécepteurs sensibles à l’ultraviolet, au bleu et au vert. Cette trichromatie, bien que différente de celle des humains, leur permet de percevoir un large spectre de couleurs.
- Types d’ommatidies : Trois types d’ommatidies (type I, II et III) qui se distinguent par la proportion de chaque type de photorécepteur. Cette organisation pourrait refléter une adaptation à la perception de différents aspects de l’environnement, comme le ciel et les fleurs.
- Rôle de l’opsine Amlop1 : L’opsine Amlop1, présente dans les photorécepteurs sensibles au vert des yeux composés (mais pas dans les ocelles), s’avère essentielle à la vision des couleurs.
- Importance pour l’apprentissage visuel : Les yeux composés sont indispensables à l’apprentissage visuel des couleurs.
En conclusion, les yeux composés, avec leur structure complexe et leur variété de photorécepteurs, sont les organes clés de la vision chromatique chez les abeilles. Ils sont indispensables à la perception des couleurs et jouent un rôle crucial dans l’apprentissage et la mémoire visuelle.
Les ocelles des abeilles
La comparaison de l’ocelle à une « mauvaise caméra » a longtemps limité la recherche sur sa fonction, car elle mettait l’accent sur la formation d’images plutôt que sur d’autres aspects de la perception visuelle.
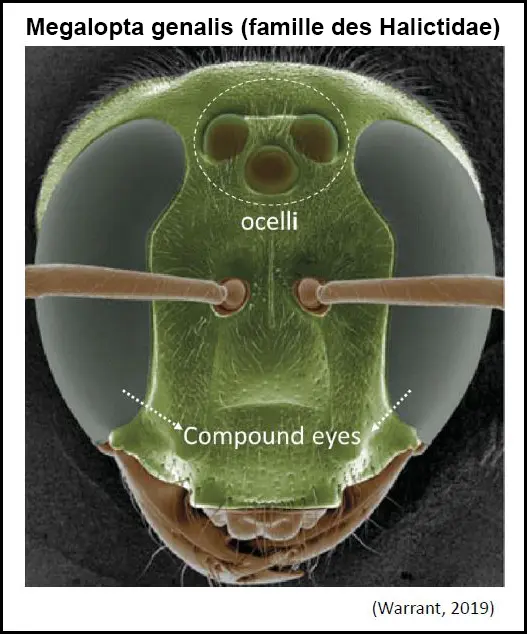
Les ocelles des abeilles, sont des yeux beaucoup plus simples disposés en triangle et situés sur le dessus de leur tête.
Ils ne perçoivent pas les détails de l’image.
Bien que petits et structurellement plus simples que les yeux composés, les ocelles jouent un rôle crucial dans la vision et le comportement des abeilles.
Les ocelles, contrairement aux yeux composés, ne sont pas impliqués dans la vision des couleurs mais plutôt dans la perception de l’intensité lumineuse (les variations d’intensité).
Leur force réside dans leur capacité à détecter rapidement les changements d’intensité lumineuse, ce qui leur permet de réagir plus vite que les yeux composés aux changements d’orientation du corps, notamment lors de mouvements rapides.
Ils agissent comme un système d’alerte précoce pour les changements d’attitude soudains, comme ceux causés par les rafales de vent.
Les ocelles jouent un rôle crucial dans la vision et le comportement des abeilles.
- Sensibilité élevée à la lumière : Les ocelles se caractérisent par leurs grandes lentilles qui les rendent plus sensibles à la lumière que les ommatidies individuelles des yeux composés.
Cette sensibilité accrue permet aux abeilles de rester actives dans des conditions de faible luminosité, par exemple au crépuscule ou à l’aube. Ce qui détermine le commencement et la fin de leurs vols de butinage. - Champ visuel large : Les ocelles des abeilles ont un large champ visuel qui couvre toute la partie dorsale et frontale de leur environnement. Cependant, chez certaines espèces d’abeilles à fourrure, comme Apis et Bombus, les poils peuvent obstruer une partie de ce champ visuel.
- Résolution spatiale faible : Bien qu’ils excellent en sensibilité à la lumière, les ocelles ont une résolution spatiale relativement faible par rapport aux yeux composés.
Cela est dû en partie à la forme irrégulière et souvent astigmate de leurs lentilles. - Perception de la lumière polarisée : La structure interne des ocelles, en particulier l’agencement des rhabdomes, les rend sensibles à la polarisation de la lumière.
Cette capacité est essentielle pour que les abeilles puissent utiliser le schéma de polarisation du ciel comme boussole pour la navigation. - Détection de l’horizon et contrôle du vol : On pense que les ocelles jouent un rôle important dans la détection de l’horizon, ce qui est crucial pour maintenir une orientation stable pendant le vol.
Cette fonction est assurée par la rétine dorsale de l’ocelle, qui perçoit l’horizon. - Influence de l’intensité lumineuse sur la taille des ocelles : Le diamètre des ocelles est corrélé au régime lumineux dans lequel les abeilles volent.
Les espèces d’abeilles qui volent dans des conditions de faible luminosité ont tendance à avoir des ocelles plus grands, ce qui souligne l’importance de la sensibilité à la lumière dans ces environnements. - Dimorphisme sexuel : Chez certaines espèces d’abeilles, la taille des ocelles peut varier entre les sexes.
Par exemple, chez de nombreuses espèces d’abeilles mellifères, les mâles ont des ocelles légèrement plus grands que les ouvrières, probablement en raison de leur rôle dans la détection des reines en vol. - Variations et adaptations spécifiques aux espèces : Bien que les ocelles partagent une structure de base chez les abeilles, il existe des variations et des adaptations spécifiques aux espèces.
Par exemple, les ocelles des abeilles euglossines (abeilles à orchidées de la famille des Apidae) sont uniques en ce sens que leur rétine dorsale se trouve dans la zone focale de la lentille, ce qui leur confère une résolution spatiale plus élevée de la scène qui se trouve devant elles.
En résumé, les ocelles sont des organes visuels essentiels pour les abeilles, qui contribuent à diverses fonctions, notamment la sensibilité à la lumière, la détection de l’horizon, la perception de la lumière polarisée et le contrôle du vol.
Bien que leur résolution spatiale soit inférieure à celle des yeux composés, leur sensibilité élevée à la lumière et leur large champ visuel en font des composants précieux du système visuel des abeilles, leur permettant de naviguer, de s’orienter et de rechercher de la nourriture efficacement dans divers environnements lumineux.
Les variations et adaptations spécifiques aux espèces de la structure et de la fonction des ocelles soulignent la diversité des adaptations visuelles chez les abeilles, ce qui met en évidence l’interaction complexe entre l’évolution, le comportement et l’écologie sensorielle.
Voici quelques exemples de fonctions assurées par les ocelles chez les abeilles :
- Détection de l’horizon pour la stabilisation du vol.
- Contrôle de l’altitude pendant le vol.
- Déclenchement du vol au crépuscule.
- Réponses phototaxiques (mouvement en réaction à la lumière).
- Obtention d’informations sur la direction à partir de repères célestes, comme la lumière polarisée du ciel.
Il est important de noter que les ocelles ne contribuent pas à la vision des couleurs.
Bien qu’ils possèdent des photorécepteurs sensibles à différentes longueurs d’onde, notamment dans l’ultraviolet et le vert, ces informations ne sont pas traitées par des neurones permettant la perception des couleurs, contrairement à ce qui se passe dans les yeux composés.
L'implication du cerveau des abeilles dans la vision des couleurs
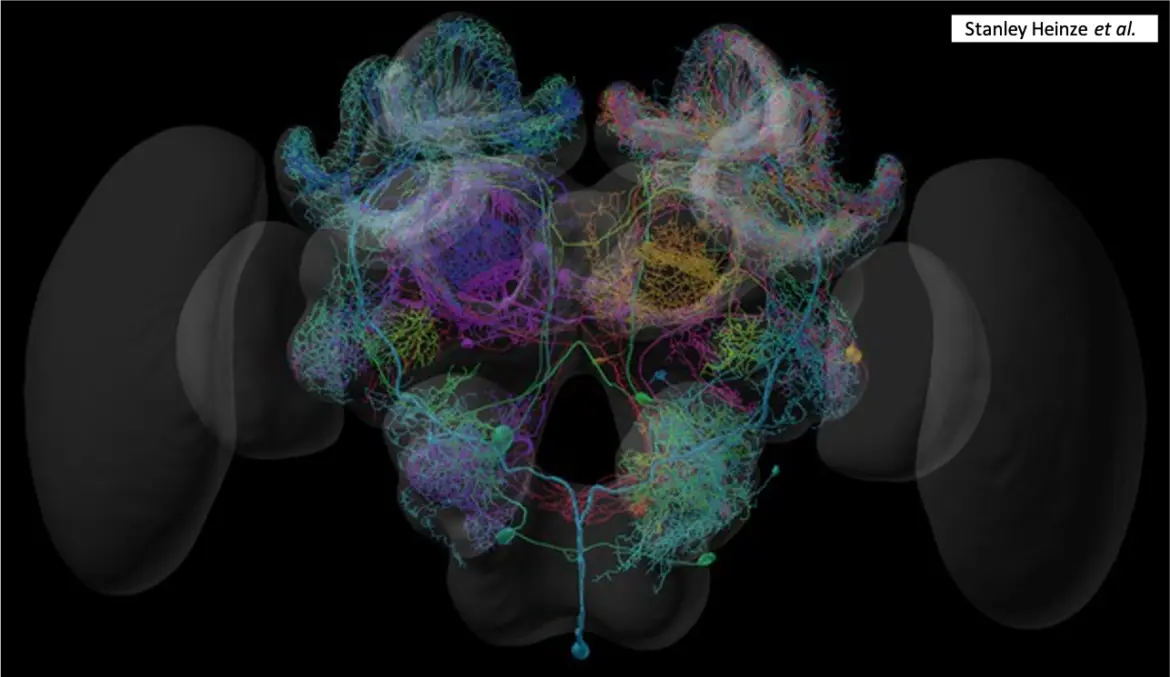
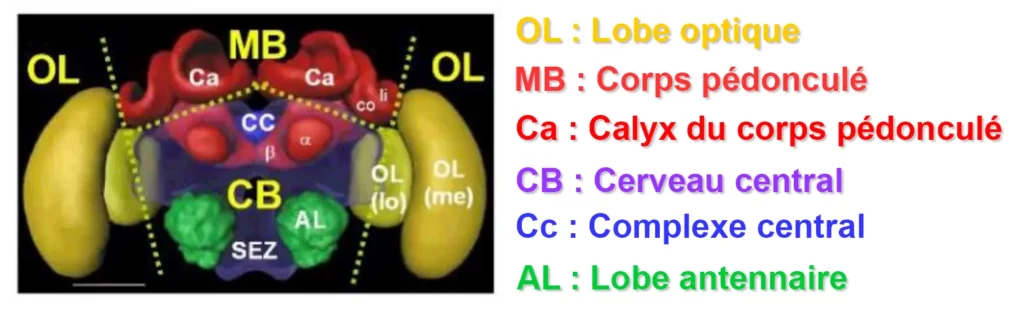
Voici les différentes régions du cerveau des abeilles impliquées dans la vision des couleurs et l’apprentissage visuel :
Lobes optiques : Les lobes optiques sont la première région du cerveau à recevoir des informations visuelles des yeux.
Ils sont responsables du traitement de l’information sur la couleur, la forme et le mouvement.
Corps pédonculés : Les corps pédonculés sont impliqués dans l’apprentissage et la mémoire, y compris l’apprentissage visuel.
Les calices sont la région d’entrée des corps pédonculés. Ils sont subdivisés en trois régions : la lèvre, le col et l’anneau basal.
Chaque région reçoit une entrée sensorielle spécifique : la lèvre, l’entrée olfactive, le col, l’entrée visuelle et l’anneau basal, l’entrée olfactive et mécanosensorielle.
Complexe central : Le complexe central est impliqué dans l’intégration sensorimotrice et le contrôle moteur.
Il reçoit également des informations visuelles des lobes optiques et on pense qu’il joue un rôle dans l’apprentissage et la mémoire visuels.
Tubercule optique antérieur : Le tubercule optique antérieur reçoit des informations des lobes optiques et serait impliqué dans le traitement des informations de couleur.
Les neurones sensibles aux couleurs des lobes optiques se projettent dans les corps pédonculés et le complexe central. Les corps pédonculés reçoivent des informations des lobes optiques ainsi que d’autres zones du cerveau, et on pense qu’ils jouent un rôle dans l’apprentissage et la mémoire associatifs.
Le complexe central reçoit des informations des lobes optiques et d’autres zones du cerveau, et on pense qu’il est impliqué dans une variété de fonctions, y compris le traitement des informations spatiales et la navigation.
Les IEG, Gènes précoces immédiats : L’apprentissage visuel induit des changements dans l’expression des gènes précoces immédiats (IEG) dans le cerveau des abeilles.
Les IEG sont un groupe de gènes qui sont rapidement exprimés en réponse à une variété de stimuli, y compris l’apprentissage et la formation de la mémoire.
L’expression des IEG est régulée à la hausse dans les corps pédonculés et les lobes optiques des abeilles qui ont appris une tâche de discrimination visuelle. Ces résultats suggèrent que ces régions du cerveau sont impliquées dans le traitement et le stockage des souvenirs visuels.
En résumé, la stimulation des couleurs chez les abeilles est le résultat d’une interaction complexe entre la physique de la lumière, la biologie des photorécepteurs et le traitement neuronal de l’information visuelle.
L’organisation spécifique des différents types de photorécepteurs et les mécanismes d’opposition chromatique au niveau du cerveau permettent aux abeilles de percevoir un monde riche en couleurs, essentiel pour leur survie et leur comportement.
Les gènes précoces immédiats (IEG) dans la recherche sur le cerveau des abeilles
Les sources présentent les IEG comme des outils précieux pour explorer l’activité neuronale chez les abeilles, notamment dans le contexte de l’apprentissage et de la mémoire visuelle.
Voici les points clés concernant les IEG et leur utilisation dans les études sur le cerveau des abeilles :
- Définition et rôle des IEG : Les IEG sont des gènes dont l’expression est rapidement activée en réponse à une stimulation neuronale, sans nécessiter de synthèse protéique. Ils agissent comme des marqueurs de l’activité neuronale, permettant d’identifier les régions du cerveau impliquées dans un processus cognitif particulier.
- IEG étudiés chez les abeilles : Trois IEG spécifiques : kakusei, Hr38 et Egr1. Ces gènes ont été précédemment associés à des activités telles que le butinage, l’orientation spatiale et la réponse à des stimuli sensoriels.
- Utilisation des IEG dans un contexte d’apprentissage visuel : des scientifiques ont utilisé l’analyse de l’expression des IEG pour étudier l’activité neuronale associée à l’apprentissage des couleurs dans un environnement de réalité virtuelle. L’objectif était d’identifier les régions du cerveau activées lors de l’apprentissage et de déterminer si les schémas d’activation diffèrent selon les conditions d’apprentissage.
- Résultats contrastés selon le type de réalité virtuelle : Les expériences ont révélé des résultats différents selon le type d’environnement de réalité virtuelle utilisé. Dans une réalité virtuelle 3D immersive, l’apprentissage réussi était corrélé à une augmentation de l’expression d’Egr1 dans les calices des corps crâniens, une région du cerveau impliquée dans la mémoire olfactive.
En revanche, dans une réalité virtuelle 2D plus restrictive, l’apprentissage entraînait une diminution de l’expression des trois IEG (kakusei, Hr38 et Egr1) dans différentes régions du cerveau, y compris les lobes optiques et les calices des corps crâniens. - Interprétations et hypothèses : Les auteurs suggèrent que la diminution de l’expression des IEG dans la réalité virtuelle 2D pourrait refléter une augmentation de l’activité inhibitrice dans certaines régions du cerveau.
Cette hypothèse est basée sur l’idée que la RV 2D, en limitant les mouvements de l’abeille et en la forçant à se concentrer sur un nombre limité de stimuli, pourrait induire un niveau de contrôle attentionnel plus élevé, nécessitant une inhibition accrue de l’activité neuronale non pertinente. - Défis techniques et perspectives : La combinaison de l’analyse des IEG avec d’autres techniques, telles que l’imagerie calcique ou l’électrophysiologie, pourrait fournir une image plus complète de l’activité neuronale associée à l’apprentissage et à la mémoire visuelle chez les abeilles.
En résumé, les IEG constituent un outil prometteur pour étudier les bases neuronales de la vision et de l’apprentissage chez les abeilles.
Quelle est l'influence de la lumière sur le comportement des abeilles ?
La plupart des espèces d’abeilles sont diurnes, ce qui signifie qu’elles sont actives pendant la journée.
Limitées à la journée pour la recherche de nourriture et autres activités effectuées à l’extérieur du nid.
Cette limitation pourrait être due à des températures plus basses ou à de faibles intensités lumineuses la nuit, ou aux deux.
Cela est aussi probablement dû au fait que leurs yeux sont adaptés aux niveaux de lumière plus élevés présents pendant la journée.
La taille des lentilles des yeux des abeilles (les facettes des yeux composés et les ocelles) détermine l’intensité lumineuse la plus faible à laquelle une espèce peut être active.
Les abeilles dotées de facettes et d’ocelles plus grands peuvent voler dans des conditions de faible luminosité, comme au crépuscule ou à l’aube.
Certaines espèces d’abeilles ont évolué pour devenir nocturnes.
Ces abeilles ont généralement des yeux plus grands avec de plus grandes facettes, ce qui leur permet de capter davantage de lumière dans des conditions de faible luminosité.
Les abeilles nocturnes s’appuient également sur d’autres adaptations sensorielles, telles que l’odorat, pour naviguer et trouver de la nourriture dans l’obscurité.
Bien que les différences de température entre le jour et la nuit puissent être marquées dans les climats tempérés et arctiques et dans les habitats secs, elles sont beaucoup plus faibles et moins susceptibles de limiter le vol des abeilles dans les zones humides et tropicales.
Ainsi, les abeilles sont généralement plus limitées par les intensités lumineuses que par les basses températures.
Il n’est donc pas surprenant que le diamètre de la lentille des deux types d’yeux d’abeilles, les lentilles des facettes des yeux composés et les lentilles ocellaires, et donc leur sensibilité, dictent la plus faible intensité lumineuse dans laquelle une espèce peut être active.
Chez les abeilles qui utilisent des repères visuels pour la navigation, la distance de vol peut au moins partiellement dépendre des capacités visuelles. Étant donné que les abeilles en vol peuvent rarement être suivies dans la nature, les distances de recherche de nourriture ont été estimées à l’aide d’expériences de lâcher-retour au nid avec des individus marqués ou en entraînant les abeilles à se nourrir à des distances définies et en enregistrant la distance à laquelle les abeilles cessent de visiter la mangeoire.
Les abeilles, comme de nombreux animaux volants, utilisent le flux optique pour mesurer la distance parcourue en vol vers et depuis les sources de nourriture.
Le débit optique est mesuré comme la quantité de mouvement de l’image dans le champ visuel latéral de l’œil de l’abeille lorsqu’elle vole vers un but.
Outre le retour au nid (au moins chez certaines espèces) et la recherche et le retour aux fleurs, la seule tâche visuelle importante, et la plus exigeante, qu’une abeille mâle doit effectuer est de trouver une compagne.
Les mâles recherchent des partenaires sur les sites où les femelles émergent, au niveau des ressources alimentaires et sur des sites sans ressources tels que des points de repère et des voies de passage.
La vision des abeilles nocturnes
Les abeilles sont actives la nuit grâce à des adaptations spéciales :
Les abeilles nocturnes ont développé des adaptations visuelles pour les aider à voir dans des conditions de faible luminosité :
Facettes oculaires plus grandes : Cela permet à davantage de lumière de pénétrer dans l’œil, ce qui améliore la vision dans des conditions de faible luminosité.
Sommation spatiale : Cela implique que les signaux de plusieurs cellules photoréceptrices sont combinés dans le cerveau, ce qui augmente la sensibilité à la lumière au prix de la résolution spatiale.
Sommation temporelle : Cela implique que les signaux visuels sont intégrés sur une période plus longue, ce qui augmente également la sensibilité à la lumière au prix de la résolution temporelle.
Ces adaptations permettent aux abeilles nocturnes de naviguer et de rechercher de la nourriture dans l’obscurité.
Par exemple, l’abeille nocturne Megalopta genalis peut utiliser le flux optique pour contrôler son vol dans des tunnels expérimentaux, même si elle vole cinq fois plus lentement que les bourdons diurnes.
L’abeille charpentière nocturne, X. tranquebarica, est capable de se nourrir et de retourner au nid les nuits de nouvelle lune. Elle peut même utiliser la couleur dans ces conditions.
La vision des couleurs chez les abeilles – Historique
 La vision des couleurs des abeilles a été officiellement démontrée par le lauréat du prix Nobel Karl von Frisch il y a 118 ans.
La vision des couleurs des abeilles a été officiellement démontrée par le lauréat du prix Nobel Karl von Frisch il y a 118 ans.
Avant lui, plusieurs scientifiques avaient suggéré que les abeilles pouvaient voir les couleurs (4 à 6), mais n’avaient pas fourni de preuves expérimentales solides prouvant cette capacité.
Von Frisch (1886 – 1982) a montré que les abeilles peuvent être entraînées à associer des cartons de différentes couleurs à une récompense sous forme de solution de sucrose, qu’il a placée sur une table expérimentale à laquelle les butineuses venaient régulièrement.
Après des visites répétées à une couleur récompensée, il a présenté une situation de test dans laquelle le carton de couleur était placé à côté de cartons gris achromatiques de différents niveaux d’intensité. Tous les cartons avaient au-dessus d’eux une mangeoire vide.
Les résultats de von Frisch ont montré de manière concluante qu’en choisissant une couleur récompensée, les abeilles la distinguaient de différents niveaux de cartons gris achromatiques, dont certains affichaient une intensité similaire à celle de la couleur entraînée.
Il a utilisé 16 cartons de couleur variant du violet au rouge et au violet (tels que vus par les humains). Cette méthode a prouvé que les abeilles pouvaient voir la majorité de ses cartons comme des surfaces colorées, sauf dans le cas du rouge, qui a été confondu avec un carton noir.

Plus tard, Alfred Richard Wilhelm Kühn (1885 – 1968) a étendu la démonstration à la gamme ultraviolette en utilisant des lumières spectrales produites par une lampe à mercure. De cette manière, il a été démontré que les abeilles peuvent voir et distinguer les couleurs dans la gamme de 300 nm (ultraviolet) à l’orange-rougeâtre (650 nm).
Quels sont les principaux facteurs affectant la vision des abeilles
Les abeilles dépendent fortement de leur vision pour des tâches essentielles telles que la recherche de nourriture et la navigation.
Bien que les principes communs de la conception et de la fonction des yeux soient partagés entre les espèces d’abeilles, des variations spécifiques d’espèces dans ces facteurs entraînent une gamme diversifiée d’adaptations visuelles, permettant aux abeilles de prospérer dans des niches écologiques variées.
Au sein de la famille des Apidae, la vision est affectée par divers facteurs :
Taille du corps : Il existe une corrélation positive entre la taille du corps de l’abeille, la taille de ses yeux et la taille de ses facettes. Par conséquent, les espèces d’abeilles plus grandes ont tendance à avoir une résolution spatiale et une sensibilité plus élevées, ce qui signifie qu’elles peuvent percevoir des objets avec plus de détails et dans des conditions de faible luminosité par rapport aux espèces plus petites.
L’heure de l’activité de vol : Les abeilles qui volent dans des conditions de faible luminosité, comme les espèces crépusculaires ou nocturnes, ont développé des adaptations visuelles pour améliorer leur sensibilité dans de tels environnements.
Elles ont tendance à avoir des facettes oculaires plus grandes et des ocelles plus grands, ce qui permet une meilleure collecte de la lumière.
Dimorphisme sexuel : Les yeux des abeilles mâles et femelles présentent des différences notables en termes de structure et de fonctionnalité, en particulier chez les espèces où les mâles sont impliqués dans la détection des partenaires.
Les mâles ont souvent une zone aiguë dans la partie dorsale de leurs yeux, avec de plus grandes facettes et des angles interommatidiaux plus petits, optimisés pour détecter les femelles en vol sur fond de ciel.
Stratégies d’accouplement : Les adaptations visuelles sont influencées par les stratégies d’accouplement employées par différentes espèces d’abeilles.
Par exemple, les abeilles charpentières mâles, qui défendent les ressources, ont tendance à avoir des zones aiguës dorsales avec des facettes plus grandes et plus nombreuses et des angles interommatidiaux plus petits par rapport aux femelles. Ces adaptations leur permettent de détecter et d’intercepter efficacement les femelles.
Contraintes écologiques : Les conditions écologiques, telles que le type d’habitat et la disponibilité de la lumière, jouent un rôle dans la formation des adaptations visuelles.
Les espèces d’abeilles vivant dans des environnements de faible luminosité peuvent avoir développé une sensibilité accrue pour naviguer et rechercher de la nourriture efficacement
Comment la vision des abeilles est-elle liée à leur comportement ?
Défense du nid : Les abeilles sociales de la famille des apidés protègent l’entrée de leur nid contre les intrus.
Par exemple, chez l’espèce Tetragonisca angustula, les abeilles de garde planent devant le nid dans des positions stables afin de protéger la trajectoire de vol des intrus ; elles utilisent des repères visuels pour se positionner.
Les abeilles géantes (Apis dorsata) utilisent également des repères visuels pour défendre leur nid. Elles réagissent davantage aux objets qui se déplacent vers le haut qu’aux objets qui se déplacent vers le bas, probablement parce qu’elles perçoivent ces derniers comme des objets inoffensifs, comme des feuilles qui tombent.
Les abeilles gardiennes assises sur le rayon s’approchent et attaquent les objets (intrus potentiels) qui se déplacent vers la ruche.
L’identification des intrus, cependant, est également médiée chimiquement
Navigation : Lorsqu’une abeille quitte la ruche pour la première fois, elle effectue un vol d’apprentissage qui lui permet de retrouver son chemin jusqu’au nid.
Elle met à jour ces informations au fur et à mesure que les caractéristiques locales changent à proximité du nid, même au cours des périodes de butinage ultérieures.
Pendant ces vols, l’abeille apprend à reconnaître les emplacements importants en mémorisant des vues centrées sur le nid, ce qui l’aide à retourner au nid.
Ces vols ont été qualifiés de vols d’orientation, vols d’apprentissage ou de comportement de retour en arrière et d’observation.
Les ouvrières étendent la portée de ces vols pour couvrir des distances croissantes au fil du temps.
Lorsqu’elle effectue ces vols, une abeille quitte généralement le nid ou le lieu d’alimentation, se retourne pour lui faire face tout en reculant dans une série d’arcs successifs qui sont approximativement centrés sur le point d’intérêt, c’est-à-dire le nid ou le lieu d’alimentation. Avec l’expérience, l’abeille volera en ligne droite et n’effectuera des vols d’orientation que lorsque des points de repère locaux autour les points d’intérêt sont perturbés.
Recherche de nourriture : Les abeilles utilisent leur vision pour détecter les fleurs à distance. La résolution spatiale de leurs yeux est liée à la taille de leur corps ; les abeilles plus grandes ont une meilleure résolution et peuvent détecter les fleurs à de plus grandes distances.
La petite abeille sans dard Tetragonula carbonaria ne peut détecter les fleurs de l’arrière-plan que lorsqu’elles ont une extension visuelle de presque 10°, ce qui signifie qu’elle peut détecter une fleur de diamètre 1 cm à une distance de 6 cm seulement.
Une abeille européenne peut détecter la même fleur lorsqu’elle remplit 3° ou 5° de son champ visuel, donc à une distance de 12 à 18 cm, et un bourdon de taille moyenne peut détecter cette fleur à une distance de 30 cm.
La résolution spatiale des grandes abeilles charpentières est probablement encore plus élevée, mais elle n’a pas été déterminée de manière comportementale.
Les distances de recherche de nourriture sont affectées par la structure du paysage, comme l’ont montré Steffan-Dewenter et Kühn, qui ont comparé les abeilles européennes dans les prairies, les forêts, les terres arables et les zones bâties en Allemagne. La distance moyenne de recherche de nourriture estimée à partir des danses frétillantes était de 1,5 km.
Distances de recherche de nourriture estimées pour différentes espèces d’Apidae (étude de 2019)
Apis florea : > 800 m
Apis cerana : 1423 m
Apis dorsata : 10 km
Apis mellifera : > 10 km
Bombus muscorum : 125 m
Bombus lapidarius : 1750 m
Bombus terrestris : 1500 m
Xylocopa flavorufa : 6 km
Xylocopa violacea : 1200 m
Melipona quadrifasciata : 2 km
Plebeia droryana : 540 m
Vols d’accouplement des mâles : La tâche visuelle la plus exigeante pour une abeille mâle est de trouver une partenaire.
La structure des yeux des mâles suggère que les horaires des vols d’accouplement, les limites de la taille du corps et les rôles relatifs de la vision et de l’olfaction dans le comportement d’accouplement influencent la structure de leurs yeux.
Par exemple, les faux-bourdons des différentes espèces d’abeilles effectuent leurs vols d’accouplement à des moments différents de la journée, qui se chevauchent à peine.
En plus de ces stratégies spécifiques, la sensibilité des yeux des abeilles à l’intensité lumineuse a une incidence importante sur leur comportement, la grande A. dorsata étant la seule espèce utilisant la fenêtre horaire la plus sombre après le coucher du soleil (18h30-19h00 en Thaïlande, en février).
Le comportement d’accouplement variait également chez trois espèces sympatriques (dans la même zone géographique) d’abeilles charpentières qui sont diurnes (Xylocopa tenuiscapa et X. leucothorax) ou nocturnes (X. tranquebarica).
X. tenuiscapa se perchait à l’extérieur des nids au début de la saison et patrouillait ou se perchait le long des voies migratoires et à proximité des plantes à fleurs plus tard dans la saison. X. leucothorax patrouillait le long des voies migratoires, tandis que X. tranquebarica, une espèce nocturne, patrouillait à proximité des buissons à fleurs.
Les mâles perchés de X. tenuiscapa peuvent détecter les femelles volant à une distance de 20 m.
Le dimorphisme sexuel dans la vision des abeilles
Le dimorphisme sexuel, c’est-à-dire les différences entre les sexes d’une même espèce, est évident dans les yeux de nombreuses espèces d’abeilles.
Ces différences dans la structure oculaire sont liées aux différents rôles que jouent les abeilles mâles et femelles dans la reproduction, notamment en ce qui concerne les comportements d’accouplement.
Les abeilles femelles, responsables de tâches telles que la construction du nid, la recherche de nourriture et les soins aux couvains, nécessitent une vision optimisée pour la navigation, la détection des fleurs et la reconnaissance des formes.
Les mâles, quant à eux, ont pour objectif principal de localiser et de s’accoupler avec les femelles, ce qui nécessite une acuité visuelle adaptée à la détection des partenaires en vol.
Chez les bourdons, les mâles des espèces qui adoptent une stratégie de perchage, attendant les femelles dans des endroits précis, ont tendance à avoir des yeux et des facettes plus grands que les ouvrières.
En revanche, les mâles des espèces qui utilisent une stratégie de patrouille, volant sur de grandes distances à la recherche de femelles, ont des yeux de taille similaire à celle des ouvrières.
En résumé, le dimorphisme sexuel de la vision des abeilles illustre l’adaptation des organismes aux exigences spécifiques de leur rôle reproductif. Les différences dans la structure oculaire entre les mâles et les femelles reflètent les pressions sélectives distinctes auxquelles ils sont confrontés, mettant en évidence l’interaction complexe entre l’évolution, le comportement et la perception sensorielle.
Chez les espèces de bourdons et d’abeilles charpentières, qui dépendent dans une large mesure des signaux chimiques, les yeux des mâles et des femelles sont plus semblables que chez les espèces qui utilisent la détection visuelle du partenaire.
Chez les faux-bourdons A. mellifera, qui ont deux fois plus d’ommatidies dans chaque œil que les ouvrières, le champ visuel est également élargi par rapport aux femelles, car les deux yeux composés se rencontrent au niveau du bord dorsal et l’extension latérale de chaque œil est plus grande (2,5 mm contre 1 mm).
Des yeux élargis avec des zones dorsales aiguës sont également observés chez les mâles d’autres espèces d’abeilles domestiques, Apis florea, A. dorsata, A. cerana et A. andreniformis.
Les mâles des deux espèces asiatiques à nidification ouverte, A. florea et A. dorsata, ont les adaptations les plus extrêmes pour la détection du partenaire, A. dorsata possédant des ommatidies élargies et A. florea ayant des facettes plus petites mais plus nombreuses, une indication de regroupement spatial.
Un schéma similaire peut être observé chez les bourdons : les mâles des espèces qui adoptent une stratégie de perchage, par exemple B. confusus, B. melaleucus et B. niveatus, ont des yeux et des facettes plus grands que les ouvrières, tandis que ceux qui utilisent une stratégie de patrouille ont des yeux de taille similaire.
Etudes scientifiques sur la vision des abeilles
Alaux, C., Robinson, G.E., 2007. Alarm Pheromone Induces Immediate–Early Gene Expression and Slow Behavioral Response in Honey Bees
Avarguès-Weber, A., Mota, T., Giurfa, M., 2012. New vistas on honey bee vision.
Backhaus, W., Menzel, R., Kreißl, S., 1987. Multidimensional scaling of color similarity in bees.
Buatois, A., Flumian, C., Schultheiss, P., Avargues-Weber, A., Giurfa, M., 2018. Transfer of visual learning between a virtual and a real environment in honey bees : the role of active vision.
Buatois, A., Laroche, L., Lafon, G., Avargues-Weber, A., Giurfa, M., 2020. Higher-order discrimination learning by honeybees in a virtual environment.
Chittka, L., Beier, W., Hertel, H., Steinmann, E., Menzel, R., 1992. Opponent colour coding is a universal strategy to evaluate the photoreceptor inputs in Hymenoptera.
Farooqui, T., Vaessin, H., Smith, B.H., 2004. Octopamine receptors in the honeybee (Apis mellifera) brain and their disruption by RNA-mediated interference.
Gegenfurtner, K.R., Kiper, D.C., 2003. Color vision. Annual review of neuroscience 26, 181–206. Geng, H., Lafon, G., Avarguès-Weber, A., Buatois, A., Massou, I., Giurfa, M., 2022. Visual learning in a virtual reality environment upregulates immediate early gene expression in the mushroom bodies of honey bees.
Hertel, H., Maronde, U., 1987. The physiology and morphology of centrally projecting visual interneurones in the honeybee brain.
Hung, Y.-S., Ibbotson, M., 2014. Ocellar structure and neural innervation in the honeybee.
Iino, S., Shiota, Y., Nishimura, M., Asada, S., Ono, M., Kubo, T., 2020. Neural activity mapping of bumble bee (Bombus ignitus) brains during foraging flight using immediate early genes.
Kent, C.F., Issa, A., Bunting, A.C., Zayed, A., 2011. Adaptive evolution of a key gene affecting queen and worker traits in the honey bee, Apis mellifera.
Kiya, T., Kunieda, T., Kubo, T., 2008. Inducible- and constitutive-type transcript variants of kakusei , a novel non-coding immediate early gene, in the honeybee brain
Kohno, H., Kubo, T., 2018. mKast is dispensable for normal development and sexual maturation of the male European honeybee.
Lafon, G., Howard, S.R., Paffhausen, B.H., Avarguès-Weber, A., Giurfa, M., 2021. Motion cues from the background influence associative color learning of honey bees in a virtual-reality scenario.
Mota, T., Yamagata, N., Giurfa, M., Gronenberg, W., Sandoz, J.C., 2011. Neural organization and visual processing in the anterior optic tubercle of the honeybee brain.
Ribi, W., Warrant, E., Zeil, J., 2011. The organization of honeybee ocelli: Regional specializations and rhabdom arrangements.
Rusch, C., Alonso San Alberto, D., Riffell, J.A., 2021. Visuo-motor feedback modulates neural activities in the medulla of the honeybee, Apis mellifera.
Rusch, C., Roth, E., Vinauger, C., Riffell, J.A., 2017. Honeybees in a virtual reality environment learn unique combinations of colour and shape.
Sarma, M.S., Rodriguez-Zas, S.L., Hong, F., Zhong, S., Robinson, G.E., 2009. Transcriptomic Profiling of Central Nervous System Regions in Three Species of Honey Bee during Dance Communication Behavior.
Schwarz, S., Albert, L., Wystrach, A., Cheng, K., 2011a. Ocelli contribute to the encoding of celestial compass information in the Australian desert ant Melophorus bagoti
Si, A., Srinivasan, M.V., Zhang, S., 2003. Honeybee navigation: properties of the visually driven `odometer’
Ugajin, A., Uchiyama, H., Miyata, T., Sasaki, T., Yajima, S., Ono, M., 2018. Identification and initial characterization of novel neural immediate early genes possibly differentially contributing to foraging-related learning and memory processes in the honeybee
Wakakuwa, M., Kurasawa, M., Giurfa, M., Arikawa, K., 2005. Spectral heterogeneity of honeybee ommatidia.
Warrant, E., Kelber, A., Wallen, R., Wcislo, W., 2006. Ocellar optics in nocturnal and diurnal bees and wasps.
Yang, E.-C., Lin, H.-C., Hung, Y.-S., 2004. Patterns of chromatic information processing in the lobula of the honeybee, Apis mellifera
Wakakuwa, M., Kurasawa, M., Giurfa, M. & Arikawa, K. Spectral heterogeneity of honeybee ommatidia.
Schwarz, S., Albert, L., Wystrach, A. & Cheng, K. Ocelli contribute to the encoding of celestial compass information in the Australian desert ant Melophorus bagoti.
Taylor, G. J. et al. The dual function of orchid bee ocelli as revealed by X-ray microtomography.
Frasnelli, E.; Hempel de Ibarra, N.; Stewart, F. J., The dominant role of visual motion cues in bumblebee flight control revealed through virtual reality. Front. Physiol. 2018,
Ugajin, A.; Uchiyama, H.; Miyata, T.; Sasaki, T.; Yajima, S.; Ono, M., Identification and initial characterization of novel neural immediate early genes possibly differentially contributing to foraging-related learning and memory processes in the honeybee. Insect Mol Biol 2018,
Avarguès-Weber, A.; Mota, T.; Giurfa, M., New vistas on honey bee vision. Apidologie 2012
Buehlmann, C.; Wozniak, B.; Goulard, R.; Webb, B.; Graham, P.; Niven, J. E., Mushroom Bodies Are Required for Learned Visual Navigation, but Not for Innate Visual Behavior, in Ants. Curr Biol 2020,
Hempel de Ibarra, N.; Vorobyev, M.R.; Menzel, R. Mechanisms, functions and ecology of colour vision in the honeybee. J. Comp. Physiol. B 2014,
Ribi,W.A.;Warrant, E.J.; Zeil, J. The organization of honeybee ocelli: Regional specialization and rhabdom arrangements. Arthropod Struct. Dev. 2011,
Leys, R.; Hogendoorn, K. Correlated evolution of mating behaviour and morphology in large carpenter bees (Xylocopa). Apidologie 2008,
Taylor, G.J.; Ribi, W.; Bech, M.; Bodey, A.J.; Rau, C.; Steuwer, A.; Warrant, E.J.; Baird, E. The dual function of orchid bee ocelli as revealed by X-ray microtomography. Curr. Biol. 2016,
Koeniger, N.; Kurze, C.; Phiancharoen, M.; Koeniger, G. “Up” or “down” that makes the difference. How giant honeybees (Apis dorsata) see the world. PLoS ONE 2017,
Chakravarthi., A.; Rajus, S.; Kelber, A.; Dacke, M.; Baird, E. Differences in spatial resolution and contrast sensitivity of flight control in the honeybees Apis cerana and Apis mellifera. J. Exp. Biol. 2018,
Osborne, J.L.; Martin, A.P.; Carreck, N.L.; Swain, J.L.; Knight, M.E.; Goulson, D.; Hale, R.J.; Sanderson, R.A. Bumblebee flight distances in relation to the forage landscape. J. Anim. Ecol. 2008
Srinivasan, M.V. Going with the flow: A brief history of the study of the honeybee’s navigational ‘odometer’. J. Comp. Physiol. A 2014
Chittka, L.; Tautz, J. The spectral input to honeybee visual odometry. J. Exp. Biol. 2003,
